Expression Patterns of CATalase and Superoxide Dismutase (Cu/Zn-SOD) Genes under Drought Stress in Petunia Hybrida
Purpose: Current study aimed to investigate the effect of drought stress on the expression of CAT and Cu/Zn-SOD using Real-time PCR.
Materials: The factorial experiment including two Persian and non-Persian (Petuniahybrida Sinja) Petunia varieties and three levels of drought stress consist of 40% of field capacity (severe stress), 70% of field capacity (mild stress) and 100% of it (no stress) as control were used in three repliCATions. Sampling of leaves was performed at three duration times including: one day, one month and three months after drought stress. For normalization, the βactin housekeeping gene was used and the data were evaluated by relative measurement method.
Results: Drought stress modified the transcript levels of CAT and Cu/Zn-SOD genes. The expression of CAT and Cu/Zn-SOD genes, depending on the Persian or non-Persian varieties, showed different pattern for duration of stress and stress intensity. CAT and Cu/Zn-SOD genes expression were similar to control at one day after the stress. CAT expression at one month after the stress (vegetative growth) in the non-Persian variety and three months after stress (reproductive growth) in Persian variety showed significant increasing. The expression of Cu/Zn-SOD gene in reproductive growth was increased in the non-Persian variety and under severe drought stress it was raised up in the Persian variety.
Conclusion: Significant changes in gene expression occur mainly during reproductive stage. Increasing the expression of CAT and Cu/Zn-SOD genes compared to control condition indiCATe contributing the development of resistance in this phase.
Keywords: Petunia Hybrida; Drought Stress; Gene Expression; Antioxidant
Drought stress is the main cause of reduced plant growth in arid and semi-arid regions which triggers the plant to a series of responses at different levels of molecular, cellular and physiological aspect [1].
Drought stress effects on physiology, morphology, anatomy, chemical composition, tissue water content and many other metabolic pathways of plant. The effects depend on type and age of plant, stage of development, nutritional status, texture and structure of soil, irrigation and climate conditions (temperature, light, etc.) [2].
As plants cannot move, therefore adaptation mechanism has great importance to deal with various environmental stresses. In drought conditions, stomata are closed followed by increasing concentrations of abscisic acid (ABA) in mesophyll cells of leaves and subsequently concentration of CO2 is reduced in mesophilic tissue. In this case, the dark reactions of photosynthesis are stopped and products of the light reaction (ATP and NADPH) are not used. In such a situation because of lack of oxidation of NADPH molecules, the consumption of NADP+ is decreased to receive electrons. So the molecule of oxygen behaves as electron acceptor in transport chain of electrons and thus a variety of reactive oxygen species (ROS) are produced [3].
Producing of ROS is a result of unfavorable environmental conditions for plants such as high temperatures, heavy metals, air pollution, and lack of food, stress, cold, drought and salinity. Molecular oxygen resuscitation stage, with the high levels of energy input or electron transfer reactions are leading to the production of large amounts of ROS, including: O2.-, H2O2, OH. and 1O2. In plants, the formation pathways of ROS are including through leakage of electrons to O2, transfer activity of electron in chloroplast, mitochondria and plasma membrane or as a byproduct of various metabolic pathways which take place in different parts of the cell. Environmental stresses are resulting in increased production of ROS due to breakdown of cell homeostasis in plants. All of ROS are harmful to organelles in high amounts. When ROS levels are excessive, cellular defense mechanisms change into oxidative stress. Increased ROS production during environmental stress can be considered as a threat to lipid peroxidation, protein oxidation, destruction of nucleic acids, enzyme inhibition, activation of programmed cell death and ultimately cell death. The ROS, as secondary transmitters are also involved in a variety of cellular processes, including tolerance to environmental stresses. The role of ROS as signaling molecules or destroyer, depends on sensitive balance between ROS production and its inhibition. Because of the multiple roles of ROS, controlling the levels of ROS in cells is essential to avoid any oxidative damage. Harness and detoxifying high levels of ROS are performed through antioxidative system effectively containing antioxidants enzymes such as superoxide dismutase (SOD), CATalase, ascorbate peroxidase (APX), monodihydro-ascorbatereductase (MDHAR), dihydro ascorbatereductase (DHAR), glutathione reductase, guaiacol peroxidase (GPOX), glutathione -S- transferase (GST) and glutathione peroxidase (GPX) and non-enzymatic antioxidants such as glutathione (GSH) and ascorbate (soluble in water), carotenoids, tocopherols (fat-soluble) and flavonoids [4,5].
Higher accumulation of Na in MI48 tissues cause more oxidative stress which can be reflected by the expression of CATA and POX1 genes. The expression level of CATA and POX1 was higher in salt susceptible MI48 plants as compared to the tolerant CSR10 plants which suggest that MI48 plants are experiencing higher levels of ROS accumulation and they need more amount of CATalase to establish the homeostasis with the stress [6].
Metallo-enzyme of SOD is the effective and intracellular enzyme antioxidant that is present in all parts of cells and aerobic organelles, in exposure of oxidative stresses induced by ROS. A variety of environmental stresses often lead to increased ROS production and SOD is the first step in defense against the toxic effects of high levels of ROS for plant stress tolerance. SOD removes the O2-. by CATalyzing it to H2O2 and O2 [4].
CATalase is the enzyme containing tetramer iron with potential direct conversion of H2O2 to H2O and O2, so they are required for detoxifiCATion of ROS in stress condition [7].
Due to the increasing of drought problem as a serious case and the risk of severe water shortage in upcoming years, the use of ornamental plants adapted to drought mechanism is very important. Finding the different methods for adaptation plays an important role for discovering the tolerance mechanism. Todays, molecular modifiCATion for increasing the resistance to a variety of stresses, especially drought stress is important, therefore, the identifiCATion of the expression pattern of candidate genes for drought tolerance in plants can be a major step in eugenic of ornamental plants. Because of the strong relationship between tolerance to environmental stresses and changes of antioxidant enzymes in the photosynthetic plants and given the fact that the synthesis of each substance in cells is controlled by genes, so the alteration in gene expression of enzymes during the stress and the effect of antioxidant can be discovered by detection of genes responsible for the synthesis of these materials in plants.
Petuniasp, the ornamental species which can be cultured in the green spaces is one of annual, flowering plants belonging to the Solanaceae family. It is native to South America (Mexico and Argentina) with varieties suitable for planting in the green spaces. This plant is suitable for planting in summer because of its variety in shape, size and color of flowers and also due to long flowering period, it is widely grown in garden and considered as the most important and valuable seasonal plant [8]. This plant tolerates drought very well and its value and importance were discovered when the majority of seasonal plants disappeared except Petunia during drought occurred in the summer of 1944 in the United States [9].
Totally, the identifiCATion of candidate genes such as superoxide dismutase and CATalase antioxidant genes and their expression pattern in drought tolerance can be a major step in eugenic of ornamental plants. In other hand, due to relatively recognized and accessible of genetic map of Petunia, it’s widely used as an ornamental plant therefore it can be considered as great choice for genetic studies to achieve more insight into stress response in tolerant plants.
In this study, we assessed the expression of antioxidant enzymes such as CATalase and superoxide dismutase (Cu/Zn-SOD) by Real-time RT-PCR , as important enzyme involving in drought stress [10].
The seeds of two Petunia varieties, Petuniahybrida Vilm. (Persian) and Petuniahybrida Sinja. (Non-Persian) were prepared from the center of Plant Production and Agricultural Research Institute of Northern Khorasan and were planted in trays.
The seed lings were kept for 6 weeks, and then transferred into the pots at the greenhouse of Ferdowsi University of Mashhad. Suitable soil was selected for the pots in a ratio of 1: 1: 1 sand, peat and loam soil. The seedlings were transplanted into pots in factorial experiment with two varieties in three irrigation levels of 40, 70 and 100% of field capacity (sever, moderate and no stress respectively) in three repliCATion. AsPetunia plant is a drought tolerant, for planting of it in green garden, three levels of irrigation, 40, 70 and 100% of field capacity, were applied at flowering phase of Petunia. The treatments were performed for daily measurements of humidity of soil and adding water into each pot, and continued until last sampling taken.
Petunia is one of the ornamental plants, most widely used in green spaces and commonly planted in the summer, mainly influenced by the drought. Significant developmental stages are the vegetative growth stage and the flowering peak (reproductive stage; three times, in three different developmental stages (early, mid and late growth) was considered to assess more specifically the effect of stress for green spaces, therefore, Sampling was performed in three stages including: 24 hours after stress (7 weeks), one month after the growth in vegetative stage (10 weeks) and three months later in reproductive stage (18 weeks) in both domestic and non-Persian varieties of Petunia. In each stage, the leaves under flag leaves were taken and placed in liquid nitrogen. Then they were kept at -80 °C.
Total RNA was extracted from frozen leaves of samples the by column RNA extraction kit (DenaZist Asia Co.) according to the company guidelines. The concentration and quality of samples of RNA were determined by Nanodrop (spectrophotometer Nanodrop 2000). cDNA synthesized by cDNA synthesis kit (Vivantis Company) according to the manufacturer’s instructions.
mRNA sequence of superoxide dismutase gene (Cu/Zn-SOD) (X14352.1) and CATalase gene (AY726007.1) of Petuniahybrida, was extracted from the NCBI database and used to design primers by Allele ID software [10]. β actin housekeeping gene was used as internal control gene and also for normalizing the results of nucleic acid in gene expression assay [10-12]. All primers were manufactured by Korean OligoTM MACRO GENE Company (Table 1).
SYBR Green kit (Pars Toos Co.) was used for Real-time PCR reaction. The cDNAs were diluted by a ratio of 1 to 5 and then reaction components of Real - time PCR were prepared as following: 10 μl SYBER green, 1 μl of each primer, 2μL of cDNA (1000ng) sample, 6 μL of double-distilled water in total volume of 20 μl. The reaction samples were denatured for 10 min at 95 °C, 40 cycles of 15 s at 95 °C, 20 seconds at 60 °C and 30 sec at 72 °C with a final extension of 72 °C for 5 minutes.
The expression levels of Cu/Zn-SOD and CAT transgenes were determined by real-time PCR. In each sample, for each gene, biological tripliCATe and two technical repliCATes were considered. Fluorescent signals were obtained at each stage of polymerization in the CFXTM Real-Time System (C1000- Thermal Cycler), Bio-RAD Company.
β actin was used as a reference for normalization and all data were compared with the β actin gene and was measured by the internal software of the device.
Standard curves, melting and reproduction of treatments and the expression of CAT and Cu/Zn-SOD were analyzed by Bio-RAD software. The Rest, Bio-Rad, and Excel softwares were used to analyze the data. Comparison of mean data was performed by JMP statistical software at P <0.05 in each step, separately.
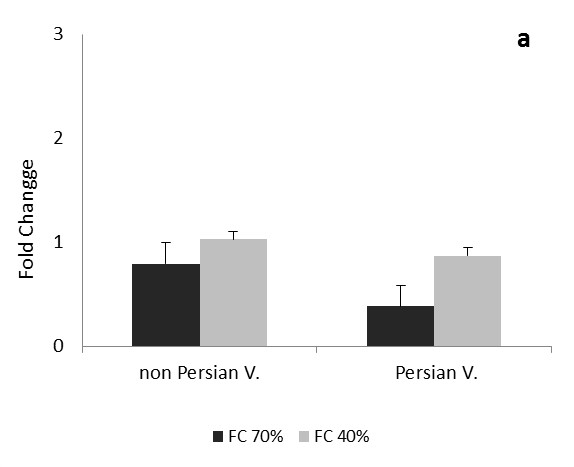
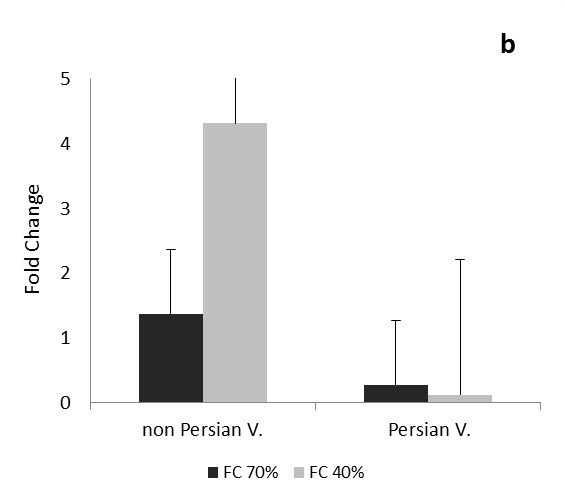
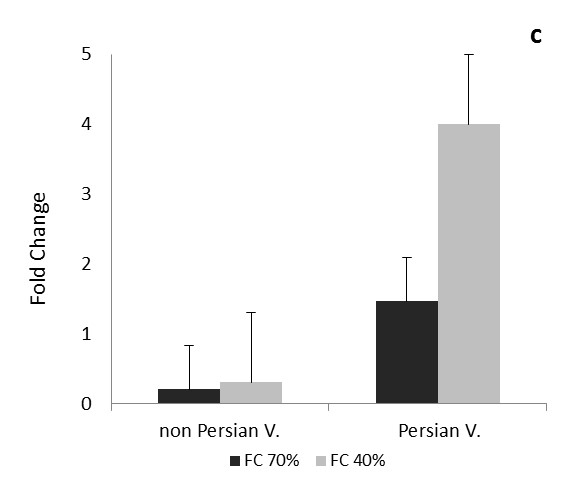
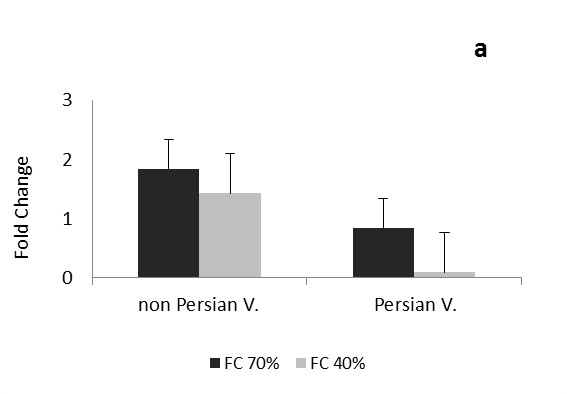
The comparison of the data in each phase and expression of CAT and Cu/Zn-SOD genes in each three times of drought, between the Persian and non-Persian varieties showed there were significant difference among three levels of field capacity (40, 70 and 100%) and the interaction of variety and field capacity 0.05 ( Figure 1 and 2).
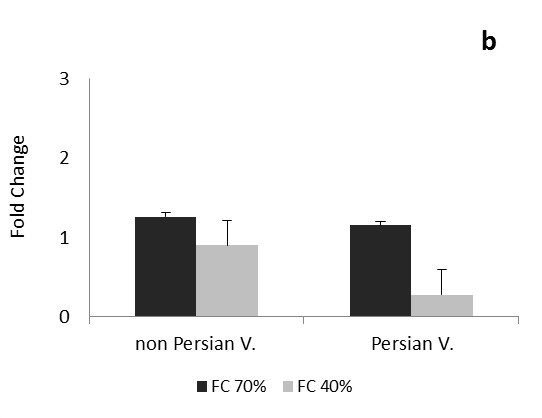
Transcription levels of CAT gene follows approximately same pattern during one dayafter drought stress in both Persian and non-Persian varieties (Figure 1). In mild stress, transcription levels of CAT gene decreased, and this was significant in Persian variety. In severe stress, CAT gene transcription level was reduced in Persian variety, but slightly increased for the Petuniahybrida Sinja., and there was no significant difference compared to the control. The expression of CAT gene increased at this time (one day after drought stress) under severe stress than mild stress, indiCATes that with increasing stress in the early stages of growth, gene expression levels of CATalase antioxidant enzyme has also increased (Figure 1a). As it is observed in Figure 1b during one month after drought stress, two varieties of Persian and Petuniahybrida Sinja (Figure 1). acted reversely and the transcription level of the CAT gene during the process of growing increased under mild and severe stress in non-Persian variety, and it was very significant in severe stress; however the transcription level of CAT gene in Persian variety showed reduction under mild and severe stress and that was very significant.
Transcription level of CAT gene decreased in non-Persian variety under both mild and severe stress at three months after drought stress, during reproductive stage and it was very significant. Persian varied showed increase for CAT gene transcript levels in both tensions and it was very significant under severe stress (Figure 1c).
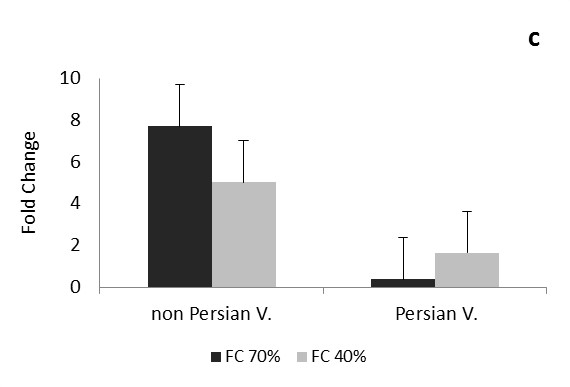
After one day of drought stress, the transcription level of Cu/Zn-SOD gene increased under mild and severe stresses in Petuniahybrida Sinja, but in Persian it was opposite and decreased (Figure 2a).
During one month after drought in Petuniahybrida Sinja, the transcript levels of Cu/Zn-SOD gene increased a little under moderate stress but decreased under severe stress; however both stress did not show any significant difference. In Persian variety the transcript levels of Cu/Zn-SOD gene increased under moderate stress but no significant difference was observed compared to the control and in severe stress condition it was greatly reduced very significantly (Figure 2b).During three months of drought stress, the transcription level of Cu/Zn-SOD gene in non-Persian variety greatly increased under mild and severe stresses. In Persian variety, gene transcript of Cu/Zn-SOD was reduced very significantly under mild stress, but under severe stress it was increased (Figure 2c).
Photorespiration increases due to result of water restrictions, stomatal closure and prevention of CO2 absorption, subsequently because of restrictions of CO2; the formation of H2O2 is increased and finally affects the expression of CAT gene. According to the previous studies, expression of CAT gene was reduced in wheat under drought. CAT gene expression under drought stress, has not changed significantly compared to the control in two tolerant and C3 sensitive genotypes of Prairie Junegrass and Kentucky bluegrass [13,14].
In this experiment, transcription of CAT gene of Petunia was changed in all three times after stress. Alteration of CAT gene expression was higher in vegetative and reproductive stages and during these periods, the action of both Petuniahybrida Sinja and Persian varieties was opposite of each other and under severe drought stress, the expression level of gene in Petuniahybrida Sinja. variety was increased in vegetative and in reproductive stage, the expression of Persian variety has increased.CAT gene transcription levels was significantly reduced at one month after drought stress (vegetative stage) in Persian variety and three months (reproductive stage) in Petuniahybrida Sinja. In Petuniahybrida Sinja. variety, the CAT gene transcript levels increased significantly in severe stress at one month after drought stress, while in Persian variety, overexpression was seen three months after severe stress, which shows the increased level of CAT gene transcription in the vegetative stage for non-Persian and in reproductive stage for Persian variety. Results showed that, the overexpression of CAT gene in Petuniahybrida Sinja. In growth stage and reversely, in the Persian variety in reproductive stage suggest that drought stress affects the Petunia in vegetative and reproductive stages more than the other early stages of growth. With increasing the age, the tolerance of Persian variety would be higher than Petuniahybrida Sinja. Species because of overexpression of CAT gene.
The higher maintenance of leaf RWC in CSG8962 (Karnal Chana 1) salt tolerant variety at 120 mM salt stress show its greater efficiency of water absorption and avoiding tissue dehydration. It may also acclimatize the crop growth under salt stress, either down playing water loss by reducing above-ground transpiration or enhancing water uptake by partitioning more assimilates towards the root system [15].
Superoxide dismutase enzyme which is product of Cu/Zn-SOD gene is the first defense line against reactive oxygen species induced by abiotic stresses. According to previous studies, the expression of Cu/Zn-SOD was decreased in leaves of Kentucky bluegrass under drought stress [13]. The level of expression of Cu/Zn-SOD did not change in sensitive and tolerant genotypes of Prairie Junegrass under drought condition [14]. In Pea, gene expression of Cu/Zn-SOD was reduced after drought stress compared to control [16].
As the Cu/Zn-SOD is an important gene in drought tension, expression change in Persian and Petuniahybrida Sinja. were expected under drought stress. Increasing expression of Cu/Zn-SOD in non-Persian variety and decreasing in Persian variety, one day after the drought indiCATes that non-Persian variety tolerates the stress condition more than Persian at early stages of drought; therefore it is leading to increased tolerance to oxidative stress induced in non-Persian variety. In Petuniahybrida Sinja. variety one day after severe drought stress, the expression of Cu/Zn-SOD was less than moderate drought, suggesting that overexpression of plant under severe stress, but stress intensity has affected the gene expression and made it much less. In Petuniahybrida Sinja. Variety, for all three times the expression of Cu/Zn-SOD increased initially in mild stress, but it was increased slightly in severe stress than moderate stress; this shows the value of Cu/Zn-SOD is reduced in greater stress rather than mild drought and its ability against oxidative stress has become more less. In Persian variety, one month after the mild drought stress, the expression of Cu / Zn-SOD was not significantly different from control, but it was decreased in severe stress, which indiCATes that Persian variety tolerates the mild stress in vegetative stage, but with increasing the stress, tolerance decreases. During the three months after the stress (reproductive stage) in Persian variety, unlike two previous times, the expression increased in severe tension, which may indiCATes that Persian variety could deal more with oxidative and drought stresses at the reproductive stage by increasing the age. In non-Persian variety the expression of Cu/Zn-SOD increased significantly at the reproductive stage, indiCATing this variety is more able to withstand the tension than the two previous tension time and also compared to the Persian variety. Variation of expression of Cu/Zn-SOD in Persian variety is lower than Petuniahybrida Sinja, suggests that the Petuniahybrida Sinja. Possibly better tolerates the drought stress at reproductive stage.
Superoxide dismutase gene is product of Cu/Zn-SOD and CATalase enzyme is made by CAT gene. Superoxide dismutase is the first defense line against reactive oxygen species induced by abiotic stresses and CATalase enzyme is the initial one that inhibits the overproduction of H2O2 under abiotic stresses. Therefore, in this study the reaction of the first lines of defense of Petuniahybrida was investigated in antioxidant defense system. Under drought stress, the expression of Cu/Zn-SOD and CAT genes changed significantly. Generally at the early stages of stress, one day after drought, the expression of both CAT and Cu/Zn-SOD genes remain at baseline and it was similar to the control plants (100% field capacity) or it is affected by the stress and it was reduced reduction. The same pattern was observed after adapting to the stress conditions at the vegetative stage, only in the Petuniahybrida Sinja. Significant changes in gene expression are observed mainly in reproductive phase and the overexpression of these genes are contributing to develop the resistance compared to control.
It is recommended to assess the physiological and morphological characteristics of Petuniahybrida under drought stress to adapt the molecular characteristics with the physiological and morphological traits. For the next examinations, also it is suggested to study the gene expression of ascorbate peroxidase, glutathione reductase under drought in this plant.
In present study, it would be better to investigate another drought gene involved in Petuniahybrida under drought stress to adapt the molecular and morphological characteristics; but this is required more fund for purchasing experimental kit.